For Week 9, I ran the RTq-PCR process on the samples I had prepared the week before. Everything went well - I actually micropipetted effectively. However, there were a couple hiccups.
Even before we started, we noticed from our assessment of the data from the spectrophotometer that we didn't have a very large concentration of RNA in most of our samples. That meant we would have to make the RTq-PCR procedure count when we did it - we would have to amplify the little we had as best as we could. That was only a minor setback though - the RTq-PCR process did amplify our samples well enough that we could see proper results.
That, however, brought us to the second problem.
When we run the RTq-PCR process, we run them with two primers, Actin and RdL. For anyone who doesn't know, primers are used to start the process of building the complementary strand. The way Rtq-PCR works is that you use enzymes to build a complementary strand of DNA for the RNA, then split the two strands apart and amplify the DNA (any general biology textbook will explain that amplification works by building a complementary DNA strand, and then splitting and building again, and so forth). But to start the process of building the complementary strand, you need primers for the enzymes to have a starting reference point, so to say:
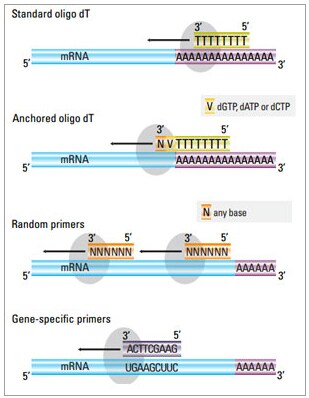
(Pic Credits: https://www.thermofisher.com/us/en/home/brands/thermo-scientific/molecular-biology/molecular-biology-learning-center/molecular-biology-resource-library/basic-principles-rt-qpcr.html)
Now, we use two primers, Actin and RdL. The Actin primer would be the reference point to build the DNA corresponding to the "Actin" RNA, and the RdL primer would be the reference point to build the DNA corresponding to the "RdL" RNA. There should be more RNA corresponding to Actin than RNA corresponding to RdL proteins from the beginning, since Actin is a really common protein crucial to cellular structure, meaning that when we run the RTq-PCR process, it should take fewer cycles to completely amplify the genetic material corresponding to Actin compared to the material corresponding to the RdL proteins. For all of our samples we ran, we compare the amplification plots to make sure this is the case. The purpose of using the Actin primer is thus to make sure we have a "control."

(Pic Credits: https://www.vet.k-state.edu/education/anatomy-physiology/faculty-staff/faculty/marcus/quan.html)
Notice in the pic above that the red plot didn't take as many cycles to reach the highest point compared to the blue plots, so there must have been more of the genetic material corresponding to the red plot. This is the kind of comparison we make. Obviously, this is just a random graph I took from online, but you get the message.
So anyway, we would make sure that everything looks normal for each sample, and then, we would just compare the RdL curves across every sample. However, the problem we ran into was that for a lot of our samples, we didn't actually get a clearly defined difference between the RdL and Actin curves. Dr. Sinakevitch fiddled around with the data, however, and we did manage to make some sense out of our data for presentation. Unfortunately, there was nothing that could be done to make the graphical data look better, so we can't use it. Probably, our plots looked really weird either because our bees were unhealthy, or I did something wrong while doing the procedure.
Kind of a bummer going into Week 10, but at least we did get conclusions for presentation.
Week 10 was a throwback to my very first days in the lab. For Week 10, I went back to doing behavioral work with PER (Proboscis Extension Reflex) conditioning. Just to review, conditioning involves using the Proboscis Extension Reflex, which is a reflex in honeybees that automatically extends their proboscises whenever their antennae are touched with a source of sugar. We condition bees to associate certain scents with sugar, so that they will extend their proboscises without being touched. The behavioral/molecular biology experiment involves conditioning bees injected with dsiRNA, Scrambled RNA, or no RNA at all (see previous posts if you don't know what those are), and then running RNA Isolation and RTq-PCR to determine the levels of RdL RNA for the bees.
During Week 10, I assisted Giulia, the undergraduate in this project, with the conditioning experiments. I caught and harnessed the bees she would use for conditioning trials, and Dr. Sinakevitch did the injections. We conditioned non-injected and dsiRNA-injected bees. For some reason, a lot of our bees died before we could condition them (lots of cleaning to do!). It was a big setback, and quite depressing to have so many deaths, but we had enough bees to at least condition a few of them. We then extracted the brains from those bees, to run the RNA Isolation and RTq-PCR process again.
Next week, I'll probably be doing behavioral work again with Giulia. We may go back to neuroanatomical work and surgeries, but it's unlikely - we got enough conclusions from our neuroanatomical work before.
I'm excited going into Weeks 11 and 12. Hope my last two weeks are fruitful!















